日本人扁桃組織由来EBウイルス株の解析によるウイルスゲノム反復配列の個人差
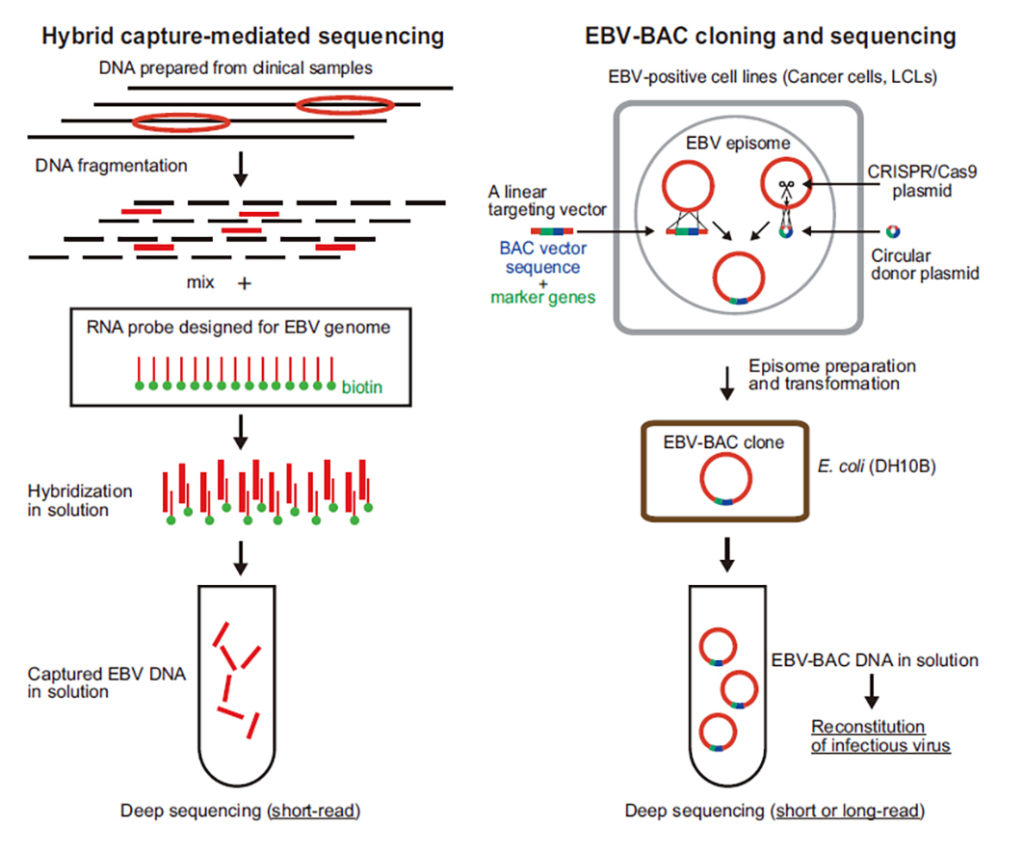
近年、英国のグループが先導して世界各地のEBウイルス臨床株の解析が進み、ウイルスゲノムには想像以上の多様性が存在することが明らかになりました。中国・香港ではEBウイルス陽性で知られる上咽頭がんの発症に関与するEBウイルス株の存在を念頭においたウイルスゲノム塩基配列の解読が進んでいます。日本では、名古屋大学のグループが日本人の伝染性単核症やEBウイルス陽性T/NKリンパ腫のEBウイルス株のウイルスゲノム塩基配列を報告しました。これらの研究はいずれもEBウイルスDNAに相補的なRNAプローブを用いてウイルスDNAを捕捉・濃縮する「ハイブリッドキャプチャー法」とショートリードシークエンサーによる塩基配列決定を組み合わせて行われました。
当教室は、本学耳鼻咽喉科と共同研究を行い、上記とは全く異なる手法によりヒト扁桃組織に感染しているEBウイルス株のウイルスゲノム塩基配列を決定しました。まず手術で摘出した扁桃組織を培養し、自然に不死化したEBウイルス感染Bリンパ芽球様細胞株を得ました。この細胞株には、1)に記載した胃がん細胞の場合と同様に、環状EBウイルスゲノムが潜伏感染しています。そこで「ゲノム編集法」により環状EBウイルスゲノムDNAをBACベクターにクローン化しました。そしてクローン化したウイルスゲノムDNAを大量調製して、ロングリードシークエンサーで塩基配列決定を行いました。ロングリードシーケンサーを用いることで、ショートリードシークエンサーが苦手とするウイルスゲノム上の反復塩基配列を正確に決定できます。
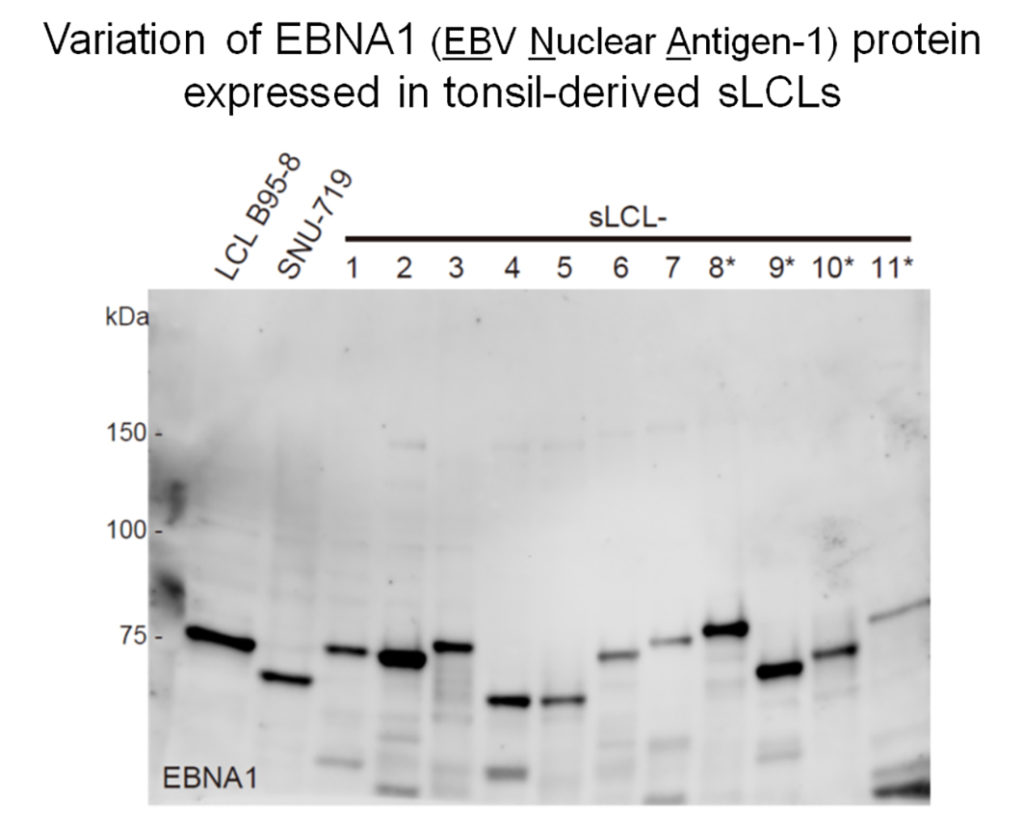
個人間での反復配列の多様性を示す一例として、EBNA1蛋白質の電気泳動上での泳動度の違いを示します。EBNA1蛋白質にはGlycine-Alanineリピートと呼ばれる繰り返し配列があり、この繰り返し配列のコピー数の違いが蛋白質の泳動度の違いとして現れます。ロングリードシークエンサーで決定したEBNA1遺伝子の反復塩基配列のウイルス株ごとのコピー数は、電気泳動上のEBNA1蛋白質の移動度とよく一致しました。